Chapter 15: Evolution on a Small Scale
15.1 Natural Selection
Natural selection is a process that leads to the adaptation of a population to both living and nonliving components of the environment.
Biotic components of the environment include competition, predation, and parasitism, which organisms respond to in order to acquire resources.
Abiotic components of the environment include weather conditions, which are dependent on temperature and precipitation.
Charles Darwin believed that species evolve over time and suggested natural selection as the mechanism for adaptation to the environment.
Darwin's hypothesis of natural selection is consistent with modern genetics, as stated in Table 15.1.
Natural selection results in the fittest individuals becoming more prevalent in a population, leading to changes over time.
The fittest individuals are those that reproduce more than others and are better adapted to the environment in most cases.
Types of Selection
Multiple alleles can produce a range of phenotypes.
Frequency distributions of phenotypes in a population often resemble bell-shaped curves.
Natural selection works to decrease detrimental phenotypes and favors those better adapted to the environment.
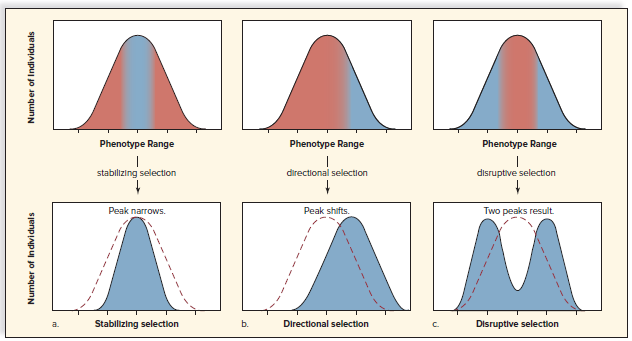
Three types of natural selection: stabilizing, directional, and disruptive.
Stabilizing selection favors intermediate phenotypes and selects against extreme phenotypes.
Directional selection favors extreme phenotypes and shifts the frequency distribution curve in that direction.
Resistance to antibiotics and insecticides is an example of directional selection.
The human struggle against malaria is an example of directional selection.
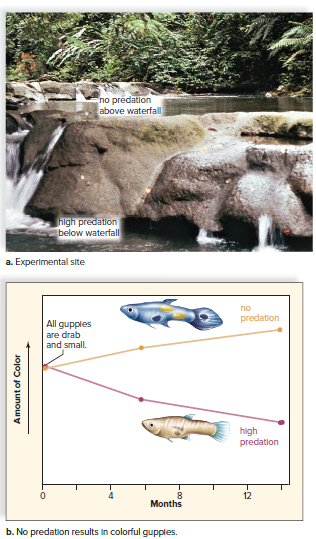
Directional selection was observed in a guppie experiment.
The environment had two areas: below the waterfall with pike and above the waterfall without pike.
Natural selection favored small and drab-colored male guppies in the lower area to avoid detection by the pike.
Male guppies moved to the area above the waterfall showed a change in phenotype towards larger, more colorful guppies.
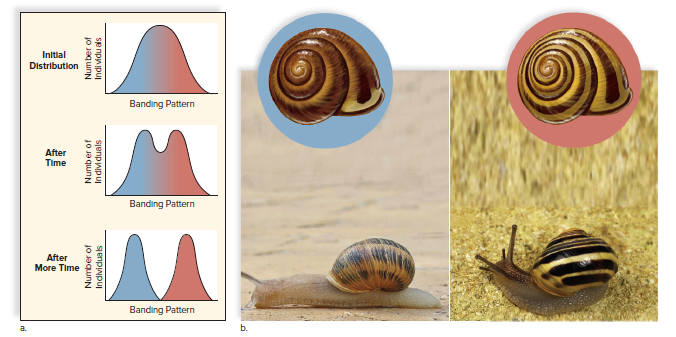
Disruptive selection favors two or more extreme phenotypes over intermediate phenotypes.
Disruptive selection favors polymorphism.
Example: British land snails found in low-vegetation areas and forests.
Thrushes feed mainly on snails with dark shells without light bands in low-vegetation areas.
Thrushes feed mainly on snails with light-banded shells in forest areas.
Two distinctly different phenotypes, each adapted to its own environment, are found in this population.
Sexual Selection
Sexual selection refers to adaptive changes in males and females that increase their ability to secure a mate.
Each sex has a different strategy for sexual selection.
Females produce few eggs, so the choice of a mate is important.
Males can father many offspring because they continuously produce sperm in great quantity.
Sexual selection in males usually results in an increased ability to compete with other males for a mate.
Sexual selection in females favors the choice of a single male with the best fitness.
Males often demonstrate their fitness by coloration or elaborate mating rituals.
By choosing a male with optimal fitness, the female increases the chances that her traits will be passed on to the next generation.
Sexual selection is considered a form of natural selection by many.

Adaptations Are Not Perfect
Natural selection does not always produce perfectly adapted organisms to their environment.
Evolution is constrained by the available variations, and each species must build upon its own evolutionary history.
The amount of variation that may be acted on by natural selection is limited.
As adaptations evolve in a species, the environment may also change.
Most adaptations provide a benefit to the species for a specific environment for a specific time.
Imperfections are common because of necessary compromises.
The success of humans is attributable to their dexterous hands, but the spine is subject to injury because the vertebrate spine did not originally evolve to stand erect.
A feature that evolves has a benefit that is worth the cost.
Maintenance of Variations
A population always has genotypic variation.
Variation is beneficial for adaptation to new environments and avoiding extinction.
Forces that promote variation are always at work: mutation, recombination, independent assortment, and fertilization.
Gene flow between populations can introduce new alleles.
Natural selection favors certain phenotypes, but other phenotypes may remain in the population at a reduced frequency.
Disruptive selection promotes polymorphism in a population.
Heterozygotes in diploid species help maintain variation by conserving recessive alleles in the population.
The Heterozygote Advantage
Only expressed alleles are subject to natural selection.
Heterozygotes can protect recessive alleles from being weeded out.
Heterozygotes can maintain the possibility of a recessive phenotype with greater fitness in a changed environment.
Balanced polymorphism occurs when natural selection maintains two different alleles of a gene at a certain ratio.
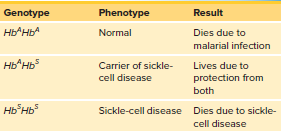
Sickle-cell disease is an example of balanced polymorphism.
HbSHbS genotype leads to sickle-cell disease and early death.
HbAHbS genotype leads to sickle-cell trait and better survival in low-oxygen environments.
HbAHbA genotype is the fittest under normal conditions.
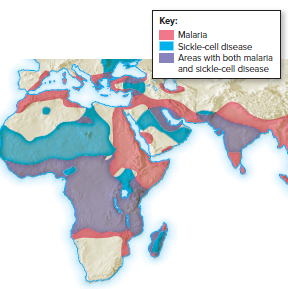
Malaria is prevalent in regions with a higher frequency of the recessive allele (HbS).
Heterozygotes (HbAHbS) are favored in malaria-prevalent environments because the parasite cannot live in their sickle-shaped red blood cells.
Homozygotes are selected against, but the recessive allele is maintained in the population.
Table 15.2 summarizes the effects of the three possible genotypes.
15.2 Microevolution
Traits can change temporarily in response to a varying environment.
Examples include:
Arctic foxes change fur color from brown to white in winter.
Dog's fur increases in thickness in cold weather.
Skin bronzing, when exposed to the sun, lasts only for a season.
These are not evolutionary changes because they are not heritable
Evolution causes a change in a heritable trait within a population, not within an individual, over many generations.
Darwin observed that populations, not individuals, evolve.
Genes interact with the environment to determine traits.
Evolution is about genetic change, specifically the change in allele frequencies in a population over time.
This type of evolution is called microevolution.
Evolution in a Genetic Context
Biologists applied genetics principles to populations in the 1930s.
They developed a way to recognize evolution and measure population changes.
In population genetics, the gene pool is made up of various alleles at all gene loci in individuals.
The gene pool is described in terms of genotype and allele frequencies.
Genotype frequency is the percentage of a specific genotype in a population.
Allele frequency represents how much a specific allele is represented in the gene pool.
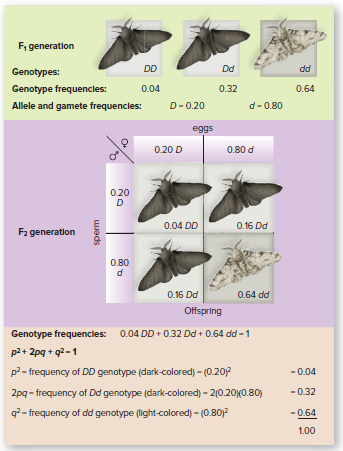
Peppered moths can be light or dark-colored.
Color is controlled by a single set of alleles: D = dark color, d = light color
In a population of peppered moths in Great Britain before pollution darkened trees:
4% were homozygous dominant (DD)
32% were heterozygous (Dd)
64% were homozygous recessive (dd)
Allele frequencies can be calculated from genotype frequencies
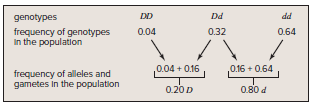

The population has a 20% frequency of the D allele and an 80% frequency of the d allele.
Gametes have a 20% chance of carrying the D allele and an 80% chance of carrying the d allele.
A punnett square can be used to calculate genotype ratios in the next generation, assuming random mating.
To produce a DD moth, both parents must contribute the D allele (20% chance each).
The probability of both events occurring is 0.20 x 0.20 = 0.04 (4%).
Therefore, 4% of the next generation should be homozygous dominant (DD).
Genotype frequencies in the next generation are the same as in the previous generation.
Sexual reproduction alone cannot bring about a change in genotype and allele frequencies.
Dominance does not cause an allele to become a common allele.
Allele frequencies of the gene pool remain at equilibrium from one generation to the next.
Recognized in 1908 by G.H. Hardy and W. Weinberg
The developed binomial equation to calculate genotype and allele frequencies
p = frequency of dominant allele, q = frequency of recessive allele
Equation: p² + 2pq + q² = 1
Five conditions must be met for mathematical relationships to remain in effect:
No mutations
No gene flow
Random mating
No genetic drift
No selection
Microevolution occurs when conditions are not met.
Deviation from Hardy-Weinberg equilibrium can measure microevolution.
These conditions are rarely if ever, met in real populations.
As a result, allele frequencies in real populations do change from one generation to the next.
This change in allele frequencies is called microevolution.
Microevolution can be detected and measured by noting the amount of deviation from Hardy-Weinberg equilibrium.
Natural selection is one of the factors that can cause microevolution.
In the case of industrial melanism, the frequency of the dark-colored phenotype increased in moth populations because the dark-colored moths were better camouflaged against the darker tree trunks that were caused by pollution.
This change in phenotype frequency was due to natural selection, and it is an example of microevolution.
Causes of Microevolution
Any conditions that change the equilibrium of alleles in a population can cause evolutionary change.
Five factors can cause a divergence from the Hardy-Weinberg equilibrium:
Genetic mutation
Gene flow
Nonrandom mating
Genetic drift
Natural selection.
Genetic Mutation
Mutations are permanent genetic changes that are the raw material for evolutionary change.
Without mutations, there can be no new variations among members of a population on which natural selection can act.
The rate of mutations is generally very low, on the order of 1 mutation per 100,000 cell divisions.
Many mutations are neutral, meaning that they are not selected for or against by natural selection.
Prokaryotes are more dependent than eukaryotes on mutations to introduce variations because they do not reproduce sexually.
All mutations that occur and result in phenotypic differences can be tested by the environment.
In sexually reproducing organisms, mutations, if recessive, do not immediately affect the phenotype.
In a changing environment, even a seemingly harmful mutation that results in a phenotypic difference can be the source of an adaptive variation.
The water flea Daphnia has a mutation that requires it to live at temperatures between 25°C and 30°C, which is adaptive only under certain environmental conditions.
Gene Flow
Gene flow is the movement of alleles among populations by migration of breeding individuals.
Gene flow can increase variation within a population by introducing novel alleles produced by mutation in another population.
Continued gene flow due to the migration of individuals makes gene pools similar and reduces the possibility of allele frequency differences among populations now and in the future.
Gene flow among populations can prevent speciation from occurring.
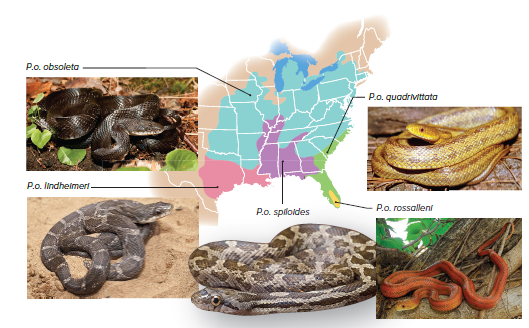
Due to gene flow, the snake populations featured in Figure 15.10 are subspecies of Pantherophis obsoleta.
These subspecies can readily interbreed when they come in contact with one another due to enough genetic similarity between the populations.
Nonrandom Mating
Random mating: individuals select mates and pair by chance, not according to their genotypes or phenotypes.
Inbreeding: mating between relatives, an example of nonrandom mating.
Inbreeding does not change allele frequencies but gradually increases the proportion of homozygotes.
Assortative mating: individuals tend to mate with those that have the same phenotype with respect to a certain characteristic.
Assortative mating causes the population to subdivide into two phenotypic classes, between which gene exchange is reduced.
Homozygotes for the gene loci that control the trait in question increase in frequency, and heterozygotes for these loci decrease in frequency.
Sexual selection favors characteristics that increase the likelihood of obtaining mates and promotes nonrandom mating.
In most species, males that compete best for access to females and/or have a phenotype that attracts females are more apt to mate and have increased fitness.
Genetic Drift
Genetic drift refers to changes in allele frequencies of a gene pool due to chance.
Allele frequencies "drift" over time and can increase or decrease depending on which members of a population die, survive, or reproduce with one another.
Genetic drift occurs in both large and small populations, but a larger population is expected to suffer less of a sampling error than a smaller population.
In a small population, random events may reduce the ability of one genotype to produce the next generation, leading to a change in allele frequencies.
Genetic drift can lead to the loss of one or more alleles, causing other alleles to become fixed in the population over time.
An experiment involving brown eye color in Drosophila flies showed that the random selection of males and females acted as a form of genetic drift, leading to the fixation of certain alleles in some populations.
Genetic drift is a random process that produces different results in different populations.
Cypress groves in California are separate populations with different phenotypes.
Phenotypes within each grove are more similar to each other than to other groves.
Environmental conditions are similar for all groves, and variations among populations are due to genetic drift.
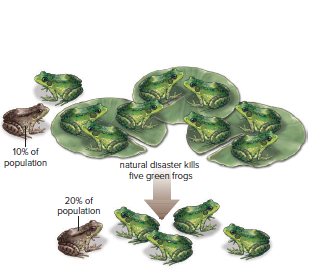
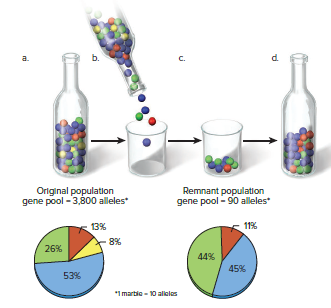
The bottleneck effect occurs when a species is subjected to near extinction, preventing most genotypes from participating in the next generation.
Cheetahs have extreme genetic similarity due to a bottleneck, causing certain alleles to be lost from the population.
The founder effect is a mechanism of genetic drift in which rare alleles occur at a higher frequency in a population isolated from the general population.
The Amish of Lancaster County, Pennsylvania, carries a recessive allele causing an unusual form of dwarfism and polydactylism at a higher frequency due to the founder effect.