0.0(0)
1.5: Nucleic acids and their functions
[Chem] Matter in our surroundings
dreissigjahriger Krieg (1618-1648)
passe compose
Timeline National Jan-June 2022
Macroevolution



Chapter 8 - Photosynthesis
Chapter 8 - Photosynthesis
- The conversion of light energy into stored chemical energy in sugars is known as photosynthesis.
- There are two types of ways to gain nutrition in general:
- Autotrophs are organisms who are able to create their own food by mainly photosynthesis.
- Heterotrophs are not able to produce their own food. They feed on autotrophs and other heterotrophs. These include decomposers. All heterotrophs depend directly or indirectly on autotrophs.
- 8.1 Photosynthesis converts light energy into the chemical energy of food:
- Chloroplasts - The sites of photosynthesis:
- Most of the plants chloroplasts are found in leaves although there are many in the stem as well. The chloroplasts are found mainly in a leaf tissue known as mesophyll.
- Inside the bilayer chloroplasts are a dense fluid known as stroma.
- Stomata are openings in the leaf by which CO2 is able to enter and O2 is able to leave.
- Thylakoids are small sacs that are hollow-ish and contain thylakoid space inside them.These are stacked up several times over into structures known as grana.
- Chlorophyll is the pigment which resides on the thylakoid membranes of the thylakoids.
- Tracking atoms through photosynthesis:
- 6CO2 + 6H2O + energy → C6H12O6 + 6O2
- This is the opposite reaction of that of cellular respiration, but is simplified to relate it to cellular respiration. In reality there are actually 12 water molecules used, but then 6 produced, so a net of 6 is written in the equation.
- The splitting of Water:
- The conclusion from this section is that the oxygen product of photosynthesis comes only from water and not carbon dioxide.
- This was tested finally by using an O_18 radioactive isotope which was put in H2O and ended up in the oxygen product only rather than glucose.
- Photosynthesis is a redox process:
- Because we know that the O in H2O becomes the oxygen as a product, the H2O is oxidized to form O2.
- The carbon in CO2 acts as a base for the building up of molecules onto it to form glucose which means that it is reduced.
- The reaction is also endergonic since it requires energy on the left side as opposed to producing energy.
- The two stages of photosynthesis - a preview:
- The light reactions are where the light is taken in and H2O is first split into O2 and H+. There is an electron shuttle like NAD+ which will be discussed later.There is also some production of ATP through the same chemiosmosis as cellular respiration in what is known as photophosphorylation.
- In the Calvin Cycle, the glucose molecule is synthesized the processes of reduction. These are sometimes called dark reactions because they occur without light, but use products of the light reactions.
8.2 The light reactions convert solar energy to chemical energy of ATP and NADPH:
- The Nature of Sunlight:
- The electromagnetic spectrum contains light and other forms of waves or energy. All EM waves travel at the speed of light but have different wavelengths. Visible light falls in between 380nm and 750nm wavelengths of the EM spectrum.
- Photons are a quantized form of light and can be thought of as discrete packets of energy. If light has a smaller wavelength, it contains more energy which means that purple light photons contain more energy than those of red light.
- Visible light is the only part of the EM spectrum that drives photosynthesis.
- Photosynthetic pigments - the light receptors:
- Pigments are materials that absorb different colors of light. When you see green, the pigment absorbs all light except for green light which it reflects.
- Spectrophotometers are devices that measure absorbance and the plot that is formed with absorption versus wavelength is known as an absorbance spectrum.
- There are three pigments in chloroplasts - chlorophyll a, chlorophyll b, and carotenoids. The action spectrum of a pigment demonstrates which color it reflects the most. The action spectrum of chlorophyll demonstrates that it does reflect green the most.
- The action spectrums of each separate pigment is less effective than that of the whole. A reflects blue-green light, B reflects olive green light.
- Carotenoids reflect orangish light and absorb some green light which means that the energy from green is not completely reflected. This pigment, however, also they act in photoprotection which dissipates excess light that is harmful to the organelle.
- Excitation of chlorophyll by light:
- When a pigment actually absorbs light, the photon energy allows for electrons to excite and raise from their ground state to higher orbitals. This is an unstable state.
- The electrons want to drop back down to the ground state and as they do they both release heat energy and light energy in what is known as fluorescence.
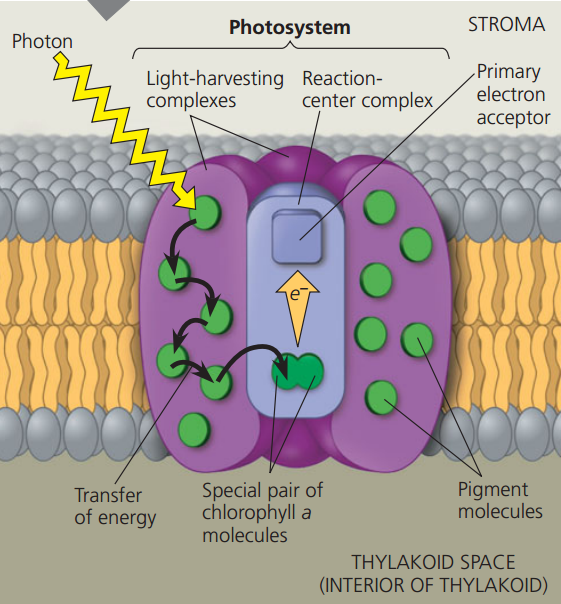
- A photosystem - a reaction-center complex associated with light-harvesting complexes:
- Photosystems are found below the pigments on the thylakoid membrane and are composed of two complexes:
- A reaction-center complex holds a special pair of chlorophyll a and a primary electron acceptor.
- A light-harvesting complex holds more pigments than the reaction center.
- Light is first absorbed by the pigments on the outside which are transferred to the photosystem. The energy is transferred until the reaction-center complex where it goes to chlorophyll a.
- The 2 chlorophyll a molecules are special in that this both boosts their energy level and also transfers the electron to the primary electron acceptor.
- There are two photosystems that are able to convert light energy and transfer it to the electron acceptor. These are Photosystem 1 (PI) and Photosystem 2 (PII). Note that PII comes before PI and is only named so because it was discovered second.
- The chlorophyll a in PI absorbs 700nm light the best and is called P700 as a result.
- The chlorophyll a in PII absorbs 680nm light the best and is called P680 as a result.
- There are slight differences, but nothing major.
- Linear Electron Flow:
- In the light reactions, there is a single direction of flow of the electron and it follows these steps:
- The photon begins in P2 and excites the chlorophyll in the light-harvesting complexes. This excitement is transferred all the way till P680 (chlorophyll a) and it raises the energy of an electron until it reaches the primary acceptor.
- The electron will later leave the P2 which leaves the P680 as a P680+ since it loses its electron. This molecule is a very strong oxidant which requires electrons. Water is split up into 2H+ and the O joins with another O that has been separated to form O2. When it is split, it gives electrons to P80+ so that it becomes P80 again. The net of this process is 2H+, 0.5O2, and electrons that go to P80+ to make it P80.
- The electron from before that has left P2 does so with Pq, a cytochrome complex, and Pc. When electrons move down the cytochrome complex, H+ moves into the thylakoid space through passive diffusion so ATP is produced through chemiosmosis. The electron travels in an electron transport chain to the P1.
- The same thing happens in P1 where P700 is excited and the electron moves to a primary acceptor forming P700+. No water is split here and so not water is formed. The electron moves down another ETC until it reaches NADP+ reductase which require 2e- and a H+ from the stroma to be an electron carrier and become NADPH.
- IMPORTANT CONCEPT NOT TAUGHT IN THE BOOK:
- The process described above is known as linear flow of electrons from P2 to P1.
- However, to maximize ATP production through photorespiration, the P1 will send back electrons to the P2 so that it can cascade down again and form ATP. This is cyclic electron flow and only starts with P1, not P2.
- A comparison of chemiosmosis:
- Basically, in mitochondria, through ATP synthase and the ETC, a lot of H+ ions are pushed out into the inter membrane space and form a pH gradient.
- In the thylakoids, the same thing happens, but within the thylakoid membrane the H+ ions are pushed. The thylakoid has a low pH and the intermembrane space of a mitochondria has a low pH.
8.3 The Calvin Cycle uses the chemical energy of ATP and NADPH to reduce CO2 to sugar:
- The Calvin Cycle is sort of like the opposite of the krebs cycle in that instead of Acetyl CoA breaking down, the process is anabolic and 3 CO2 molecules enter and are reduced by NADPH so that it can build up. This is known as carbon fixation. The result of this is not glucose but is instead glyceraldehyde 3-phosphate (G3P).
- The steps of the Calvin Cycle:
- Carbon Fixation: The 3 CO2 molecules enter one at a time and each attach to one molecule of a 5-carbon molecule known as RuBP forming an unstable 6-carbon sugar. This takes place with the enzyme rubisco which is thought to be the most abundant protein on earth.
- This then splits into two 3-carbon molecules.
- Reduction: ATP phosphorylates this sugar by adding a phosphate group and then NADPH also reduces it. It loses the phosphate group later and becomes G3P. 6 of these molecules are formed from 3 molecules of CO2 but keep in mind this is with the addition of RuBP at the beginning.
- Regeneration of RuBP: 5 of the 6 G3P molecules become 3 RuBP again so that the cycle can occur again. This process also uses ATP. The net reaction of this entire cycle is 3 CO2 and 9 ATP used to form one molecule of G3P.
- G3P is the final product of the dark reactions and is used in metabolic pathways to eventually become sugar or other macromolecules outside of photosynthesis.
- Evolution of Alternative mechanisms of carbon fixation in hot, arid climates:
- Over time, plants have developed stomata which are openings that close to prevent water loss during the day. This reduces the amount of CO2 that can be taken in and O2 that is expelled and if the stomata are closed for the majority of a day, the light is wasted and no plants can photosynthesize.
- In most plants, rubisco enables CO2 fixation to RuBP to form a 3 carbon sugar as seen before. These plants are known as C3 plants.
- What can happen that is dangerous is that CO2 becomes scarce. When this happens, Rubisco tends to be able to bind to O2 just as well as CO2 and then the calvin cycle mimics the krebs cycle. However, no ATP is produced and organic molecules are used, eventually killing the cell. This is known as photorespiration.
- C4 plants:
- C4 plants first fix CO2 into 4-carbon molecules rather 3-carbon ones.
- In the mesophyll, there is an enzyme that is able to shuttle the carbon to the bundle sheath and release the CO2 in a high enough concentration such that the calvin cycle can occur.
- This is spatial adaptation since the carbon are physically separated from the process.
- CAM plants:
- CAM plants close their stomata in the day and open it in the night to receive CO2 and O2. The CO2 is stored and fixated through Crassulacean acid metabolism (CAM).
- Then, in the day, the stomata close again but light can enter and the light reactions are able to occur.
- This is temporal adaptation since the CAM plants’ function depend on the time of day.