
Transport in Plants
Means of Transport
Diffusion:
Movement by diffusion is passive and may be from one part of the cell to the other, from cell to cell, or over short distances, say, from the intercellular spaces of the leaf to the outside.
No energy expenditure takes place.
In diffusion, molecules move in a random fashion, the net result being substances moving from regions of higher concentration to regions of lower concentration.
Diffusion is a slow process and is not dependent on a ‘living system’.
Diffusion is obvious in gases and liquids, but diffusion in solids is more likely rather than in solids.
Diffusion is very important to plants since it is the only means for gaseous movement within the plant body.
Diffusion rates are affected by the gradient of concentration, the permeability of the membrane separating them, temperature, and pressure.
Facilitated Diffusion:
The diffusion rate depends on the size of the substances; obviously smaller substances diffuse faster.
The diffusion of any substance across a membrane also depends on its solubility in lipids, the major constituent of the membrane.
Substances soluble in lipids diffuse through the membrane faster.
Substances that have a hydrophilic moiety, find it difficult to pass through the membrane; their movement has to be facilitated.
Membrane proteins provide sites at which such molecules cross the membrane.
They do not set up a concentration gradient: a concentration gradient must already be present for molecules to diffuse even if facilitated by the proteins.
This process is called facilitated diffusion.
In facilitated diffusion special proteins help move substances across membranes without the expenditure of ATP energy.
Facilitated diffusion cannot cause net transport of molecules from a low to a high concentration – this would require the input of energy.
The transport rate reaches a maximum when all of the protein transporters are being used (saturation).
Facilitated diffusion is very specific: it allows cells to select substances for uptake.
It is sensitive to inhibitors that react with protein side chains.
The proteins form channels in the membrane for molecules to pass through.
Some channels are always open; others can be controlled.
Some are large, allowing a variety of molecules to cross.
The porins are proteins that form large pores in the outer membranes of the plastids, mitochondria, and some bacteria allowing molecules up to the size of small proteins to pass through.
The transport protein then rotates and releases the molecule inside the cell, e.g., water channels – made up of eight different types of aquaporins.
Passive Symports and Antiports:
Some carriers or transport proteins allow diffusion only if two types of molecules move together.
In a symport, both molecules cross the membrane in the same direction.
In an antiport, they move in opposite directions.
When a molecule moves across a membrane independent of other molecules, the process is called uniport.
Active Transport:
Active transport uses energy to transport and pump molecules against a concentration gradient.
Active transport is carried out by specific membrane proteins.
Hence different proteins in the membrane play a major role in both active as well as passive transport.
Pumps are proteins that use energy to carry substances across the cell membrane.
These pumps can transport substances from a low concentration to a high concentration (‘uphill’ transport).
The transport rate reaches a maximum when all the protein transporters are being used or are saturated.
Like enzymes, the carrier protein is very specific in what it carries across the membrane.
These proteins are sensitive to inhibitors that react with protein side chains.
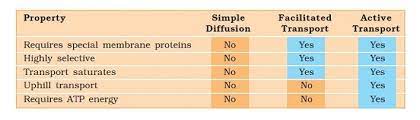
Comparison of Different Transport Processes:
Proteins in the membrane are responsible for facilitated diffusion and active transport and hence show common characteristics of being highly selective.
They are liable to saturate, respond to inhibitors, and are under hormonal regulation.
But diffusion whether facilitated or not – take place only along a gradient and does not use energy.
 Plant-Water Relation:
Plant-Water Relation:
Water is essential for all physiological activities of the plant and plays a very important role in all living organisms.
It provides the medium in which most substances are dissolved.
The protoplasm of the cells is nothing but water in which different molecules are dissolved and (several particles) suspended.
A watermelon has over 92 percent water; most herbaceous plants have only about 10 to 15 percent of their fresh weight as dry matter.
Of course, the distribution of water within a plant varies – woody parts have relatively very little water, while soft parts mostly contain it. water.
A seed may appear dry but it still has water – otherwise, it would not be alive and respiring.
Terrestrial plants take up huge amounts of water daily but most of it is lost to the air through evaporation from the leaves, i.e., transpiration.
A mature corn plant absorbs almost three liters of water in a day, while a mustard plant absorbs water equal to its own weight in about 5 hours.
Because of this high demand for water, it is not surprising that water is often the limiting factor for plant growth and productivity in both agricultural and natural environments.
Water Potential:
Water potential (Ψw ) is a concept fundamental to understanding water movement. Solute potential (Ψs ) and pressure potential (Ψp ) are the two main components that determine water potential.
Water molecules possess kinetic energy.
In liquid and gaseous form they are in random motion that is both rapid and constant.
The greater the concentration of water in a system, the greater it is kinetic energy or ‘water potential’.
Hence, it is obvious that pure water will have the greatest water potential.
If two systems containing water are in contact, random movement of water molecules will result in a net movement of water molecules from the system with higher energy to the one with lower energy.
Thus water will move from the system containing water at higher water potential to the one having low water potential.
This process of movement of substances down a gradient of free energy is called diffusion.
Water potential is denoted by the Greek symbol Psi or Ψ and is expressed in pressure units such as pascals (Pa).
By convention, the water potential of pure water at standard temperatures, which is not under any pressure, is taken to be zero.
If some solute is dissolved in pure water, the solution has fewer free water molecules, and the concentration (free energy) of water decreases, reducing its water potential.
Hence, all solutions have a lower water potential than pure water; the magnitude of this lowering due to the dissolution of a solute is called solute potential or Ψs.
Ψs is always negative.
The more the solute molecules, the lower (more negative) the Ψs.
For a solution at atmospheric pressure (water potential) Ψw = (solute potential) Ψs.
If a pressure greater than atmospheric pressure is applied to pure water or a solution, its water potential increases.
It is equivalent to pumping water from one place to another.
Pressure can build up in a plant system when water enters a plant cell due to diffusion causing pressure to build up against the cell wall, it makes the cell turgid this increases the pressure potential.
Pressure potential is usually positive, though in plants negative potential or tension in the water column in the xylem plays a major role in water transport up a stem.
Pressure potential is denoted as Ψp.
The water potential of a cell is affected by both solute and pressure potential.
The relationship between them is as follows:
Ψw = Ψs + Ψp
Osmosis:
The plant cell is surrounded by a cell membrane and a cell wall.
The cell wall is freely permeable to water and substances in solution hence is not a barrier to movement.
In plants, the cells usually contain a large central vacuole, whose contents, the vacuolar sap, contribute to the solute potential of the cell.
In plant cells, the cell membrane and the membrane of the vacuole, and the tonoplast together are important determinants of the movement of molecules in or out of the cell.
Osmosis is the term used to refer specifically to the diffusion of water across a differentially- or selectively permeable membrane.
Osmosis occurs spontaneously in response to a driving force.
The net direction and rate of osmosis depend on both the pressure gradient and concentration gradient.
Water will move from its region of higher chemical potential (or concentration) to its region of lower chemical potential until equilibrium is reached.
At equilibrium, the two chambers should have nearly the same water potential.
External pressure can be applied from the upper part of the funnel such that no water diffuses into the funnel through the membrane.
This pressure required to prevent water from diffusing is in fact, the osmotic pressure and this is the function of the solute concentration; the more the solute concentration, the greater will be the pressure required to prevent water from diffusing.
Numerically osmotic pressure is equivalent to the osmotic potential, but the sign is the opposite.
Osmotic pressure is a positive pressure applied, while the osmotic potential is negative.
Plasmolysis:
The behavior of the plant cells (or tissues) with regard to water movement depends on the surrounding solution.
If the external solution balances the osmotic pressure of the cytoplasm, it is said to be isotonic.
If the external solution is more dilute than the cytoplasm, it is hypotonic and if the external solution is more concentrated, it is hypertonic.
Cells swell in hypotonic solutions and shrink in hypertonic ones.
Plasmolysis occurs when water moves out of the cell and the cell membrane of a plant cell shrinks away from its cell wall.
This occurs when the cell (or tissue) is placed in a solution that is hypertonic (has more solutes) to the protoplasm.
Water moves out; it is first lost from the cytoplasm and then from the vacuole.
The water when drawn out of the cell through diffusion into the extracellular (outside the cell) fluid causes the protoplast to shrink away from the walls.
The cell is said to be plasmolyzed.
The movement of water occurred across the membrane moving from an area of high water potential (i.e., the cell) to an area of lower water potential outside the cell.
When the cell (or tissue) is placed in an isotonic solution, there is no net flow of water towards the inside or outside.
If the external solution balances the osmotic pressure of the cytoplasm it is said to be isotonic.
When water flows into the cell and out of the cell and is in equilibrium, the cells are said to be flaccid.
The process of plasmolysis is usually reversible.
When the cells are placed in a hypotonic solution (higher water potential or dilute solution as compared to the cytoplasm), water diffuses into the cell causing the cytoplasm to build up a pressure against the wall, which is called turgor pressure.
The pressure exerted by the protoplasts due to the entry of water against the rigid walls is called pressure potential Ψp.
Because of the rigidity of the cell wall, the cell does not rupture.
This turgor pressure is ultimately responsible for the enlargement and extension growth of cells.
Imbibition:
Imbibition is a special type of diffusion when water is absorbed by solids – colloids – causing them to increase in volume.
The classical examples of imbibition are the absorption of water by seeds and dry wood.
The pressure that is produced by the swelling of wood had been used by prehistoric man to split rocks and boulders.
Imbibition is also diffusion since water movement is along a concentration gradient; the seeds and other such materials have almost no water hence they absorb water easily.
The water potential gradient between the absorbent and the liquid imbibed is essential for imbibition.
In addition, for any substance to imbibe any liquid, the affinity between the adsorbent and the liquid is also a prerequisite.
Long Distance Transport of Water:
Long-distance transport of substances within a plant cannot be by diffusion alone.
Diffusion is a slow process.
It can account for only the short-distance movement of molecules.
In large and complex organisms, often substances have to be moved over long distances.
Sometimes the sites of production or absorption and sites of storage are too far from each other; diffusion or active transport would not suffice.
Special long-distance transport systems become necessary so as to move substances across long distances and at a much faster rate.
Water and minerals, and food are generally moved by a mass or bulk flow system.
Mass flow is the movement of substances in bulk or en masse from one point to another as a result of pressure differences between the two points.
It is a characteristic of mass flow that substances, whether in solution or in suspension, are swept along at the same pace, as in a flowing river.
This is unlike diffusion where different substances move independently depending on their concentration gradients.
Bulk flow can be achieved either through a positive hydrostatic pressure gradient (e.g., a garden hose) or a negative hydrostatic pressure gradient (e.g., suction through a straw).
How do Plants Absorb Water?
The responsibility of absorption of water and minerals is more specifically the function of the root hairs that are present in millions at the tips of the roots.
Root hairs are thin-walled slender extensions of root epidermal cells that greatly increase the surface area for absorption.
Water is absorbed along with mineral solutes, by the root hairs, purely by diffusion.
Once water is absorbed by the root hairs, it can move deeper into root layers by two distinct pathways:
The apoplast pathway is the system of adjacent cell walls that is continuous throughout the plant, except at the Casparian strips of the endodermis in the roots.
The apoplastic movement of water occurs exclusively through the intercellular spaces and the walls of the cells.
Movement through the apoplast does not involve crossing the cell membrane.
This movement is dependent on the gradient.
The apoplast does not provide any barrier to water movement and water movement is through mass flow.
As water evaporates into the intercellular spaces or the atmosphere, tension develops in the continuous stream of water in the apoplast, hence the mass flow of water occurs due to the adhesive and cohesive properties of water.
The Symplast pathway is the system of interconnected protoplasts.
Neighboring cells are connected through cytoplasmic strands that extend through plasmodesmata.
During symplastic movement, the water travels through the cells – their cytoplasm; intercellular movement is through the plasmodesmata.
Water has to enter the cells through the cell membrane, hence the movement is relatively slower.
Movement is again down a potential gradient.
Symplastic movement may be aided by cytoplasmic streaming.
You may have observed cytoplasmic streaming in cells of the Hydrilla leaf; the movement of chloroplast due to streaming is easily visible.
Most of the water flow in the roots occurs via the apoplast since the cortical cells are loosely packed, and hence offer no resistance to water movement.
However, the inner boundary of the cortex, the endodermis, is impervious to water because of a band of suberised matrix called the Casparian strip.
Water molecules are unable to penetrate the layer, so they are directed to wall regions that are not suberised, into the cells properly through the membranes.
The water then moves through the symplast and again crosses a membrane to reach the cells of the xylem.
The movement of water through the root layers is ultimately symplastic in the endodermis.
This is the only way water and other solutes can enter the vascular cylinder.
Once inside the xylem, water is again free to move between cells as well as through them.
In young roots, water enters directly into the xylem vessels and/or tracheids.
These are non-living conduits and so are parts of the apoplast.
Some plants have additional structures associated with them that help in water (and mineral) absorption.
A mycorrhiza is a symbiotic association of a fungus with a root system.
The fungal filaments form a network around the young root or penetrate the root cells.
The hyphae have a very large surface area that absorbs mineral ions and water from the soil from a much larger volume of soil that perhaps a root cannot do.
The fungus provides minerals and water to the roots, in turn, the roots provide sugars and N-containing compounds to the mycorrhizae.
Some plants have an obligate association with mycorrhizae.
For example, Pinus seeds cannot germinate and establish without the presence of mycorrhizae.
Water Movement up a Plant:
Root Pressure:
As various ions from the soil are actively transported into the vascular tissues of the roots, water follows (its potential gradient) and increases the pressure inside the xylem.
This positive pressure is called root pressure and can be responsible for pushing up water to small heights in the stem.
Effects of root pressure are also observable at night and early morning when evaporation is low, and excess water collects in the form of droplets around special openings of veins near the tip of grass blades, and leaves of many herbaceous parts.
Such water loss in its liquid phase is known as guttation.
Root pressure can, at best, only provide a modest push in the overall process of water transport.
They obviously do not play a major role in water movement up tall trees.
The greatest contribution of root pressure may be to re-establish the continuous chains of water molecules in the xylem which often break under the enormous tensions created by transpiration.
Root pressure does not account for the majority of water transport; most plants meet their need by transpiratory pull.
Transpiration pull:
Despite the absence of a heart or a circulatory system in plants, the upward flow of water through the xylem in plants can achieve fairly high rates, up to 15 meters per hour.
Most researchers agree that water is mainly ‘pulled’ through the plant and that the driving force for this process is transpiration from the leaves.
This is referred to as the cohesion-tension-transpiration pull model of water transport.
Water is transient in plants.
Less than 1 percent of the water reaching the leaves is used in photosynthesis and plant growth.
Most of it is lost through the stomata in the leaves.
This water loss is known as transpiration.
Transpiration:
Transpiration is the evaporative loss of water by plants. It occurs mainly through stomata.
Besides the loss of water vapor in transpiration, the exchange of oxygen and carbon dioxide in the leaf also occurs through these stomata.
Normally stomata are open in the daytime and close during the night.
The immediate cause of the opening or closing of stomata is a change in the turgidity of the guard cells.
The inner wall of each guard cell, towards the pore or stomatal aperture, is thick and elastic.
When turgidity increases within the two guard cells flanking each stomatal aperture or pore, the thin outer walls bulge out and force the inner walls into a crescent shape.
The opening of the stoma is also aided due to the orientation of the microfibrils in the cell walls of the guard cells.
Cellulose microfibrils are oriented radially rather than longitudinally making it easier for the stoma to open.
When the guard cells lose turgor, due to water loss (or water stress) the elastic inner walls regain their original shape, the guard cells become flaccid and the stoma closes.
Usually, the lower surface of a dorsiventral (often dicotyledonous) leaf has a greater number of stomata while in an isobilateral (often monocotyledonous) leaf they are about equal on both surfaces.
Transpiration is affected by several external factors: temperature, light, humidity, and wind speed.
Plant factors that affect transpiration include the number and distribution of stomata, percent of open stomata, water status of the plant, canopy structure, etc
The transpiration-driven ascent of xylem sap depends mainly on the following physical properties of water:
Cohesion: Mutual attraction between water molecules.
Adhesion: attraction of water molecules to polar surfaces (such as the surface of tracheary elements).
Surface Tension: water molecules are attracted to each other in the liquid phase more than to water in the gas phase.
These properties give water high tensile strength, i.e., an ability to resist a pulling force, and high capillarity, i.e., the ability to rise in thin tubes.
In plants, capillarity is aided by the small diameter of the tracheary elements – the tracheids and vessel elements.
The process of photosynthesis requires water.
The system of xylem vessels from the root to the leaf vein can supply the needed water.
As water evaporates through the stomata, since the thin film of water over the cells is continuous, it results in the pulling of water, molecule by molecule, into the leaf from the xylem.
Also, because of the lower concentration of water vapor in the atmosphere as compared to the substomatal cavity and intercellular spaces, water diffuses into the surrounding air.
This creates a ‘pull’.
Measurements reveal that the forces generated by transpiration can create pressures sufficient to lift a xylem-sized column of water over 130 meters high.
Transpiration and Photosynthesis – a Compromise:
Transpiration has more than one purpose; it
Creates transpiration pull for absorption and transport of plants.
Supplies water for photosynthesis.
Cools leaf surfaces, sometimes 10 to 15 degrees, by evaporative cooling.
Maintains the shape and structure of the plants by keeping cells turgid.
Transports minerals from the soil to all parts of the plan.
An actively photosynthesizing plant has an insatiable need for water.
Photosynthesis is limited by available water which can be swiftly depleted by transpiration.
The humidity of rainforests is largely due to this vast cycling of water from root to leaf to the atmosphere and back to the soil.
The evolution of the C4 photosynthetic system is probably one of the strategies for maximizing the availability of CO2 while minimizing water loss.
C4 plants are twice as efficient as C3 plants in terms of fixing carbon dioxide (making sugar).
However, a C4 plant loses only half as much water as a C3 plant for the same amount of CO2 fixed.
Uptake and Transport of Mineral Nutrients:
Plants obtain their carbon and most of their oxygen from CO2 in the atmosphere.
However, their remaining nutritional requirements are obtained from water and minerals in the soil.
Uptake of Mineral Ions:
Unlike water, all minerals cannot be passively absorbed by the roots.
Two factors account for this:
Minerals are present in the soil as charged particles (ions) that cannot move across cell membranes.
The concentration of minerals in the soil is usually lower than the concentration of minerals in the root.
Therefore, most minerals must enter the root by active absorption into the cytoplasm of epidermal cells.
This needs energy in the form of ATP.
The active uptake of ions is partly responsible for the water potential gradient in roots, and therefore for the uptake of water by osmosis.
Some ions also move into the epidermal cells passively.
Ions are absorbed from the soil by both passive and active transport.
Specific proteins in the membranes of root hair cells actively pump ions from the soil into the cytoplasms of the epidermal cells.
Like all cells, the endodermal cells have many transport proteins embedded in their plasma membrane; they let some solutes cross the membrane, but not others.
Transport proteins of endodermal cells are control points, where a plant adjusts the quantity and types of solutes that reach the xylem.
Note that the root endodermis because of the layer of suberin has the ability to actively transport ions in one direction only.
Translocation of Mineral Ions:
After the ions have reached the xylem through active or passive uptake, or a combination of the two, their further transport up the stem to all parts of the plant is through the transpiration stream.
The chief sinks for the mineral elements are the growing regions of the plant, such as the apical and lateral meristems, young leaves, developing flowers, fruits and seeds, and the storage organs.
Unloading of mineral ions occurs at the fine vein endings through diffusion and active uptake by these cells.
Mineral ions are frequently remobilized, particularly from older, senescing parts.
Older dying leaves export much of their mineral content to younger leaves.
Similarly, before leaves fall in deciduous plants, minerals are removed from other parts.
Elements most readily mobilized are phosphorus, sulfur, nitrogen, and potassium.
Some elements that are structural components like calcium are not remobilized.
An analysis of the xylem exudates shows that though some of the nitrogen travels as inorganic ions, much of it is carried in the organic form as amino acids and related compounds.
Similarly, small amounts of P and S are carried as organic compounds.
In addition, a small amount of exchange of materials does take place between the xylem and phloem.
Hence, it is not that we can clearly make a distinction and say categorically that xylem transports only inorganic nutrients while phloem transports only organic materials, as was traditionally believed.
Phloem Transport: Flow from Source to Sink.
Food, primarily sucrose, is transported by the vascular tissue phloem from a source to a sink.
Usually, the source is understood to be that part of the plant which synthesizes the food, i.e., the leaf, and sink, the part that needs or stores the food.
But, the source and sink may be reversed depending on the season, or the plant’s needs.
Sugar stored in roots may be mobilized to become a source of food in the early spring when the buds of trees, act as sinks; they need energy for the growth and development of the photosynthetic apparatus.
Since the source-sink relationship is variable, the direction of movement in the phloem can be upwards or downwards, i.e., bi-directional.
This contrasts with that of the xylem where the movement is always unidirectional, i.e., upwards.
Hence, unlike the one-way flow of water in transpiration, food in phloem sap can be transported in any required direction so long as there is a source of sugar and a sink able to use, store or remove the sugar.
Phloem sap is mainly water and sucrose, but other sugars, hormones, and amino acids are also transported or translocated through the phloem.
The Pressure Flow or Mass Flow Hypothesis:
The accepted mechanism used for the translocation of sugars from source to sink is called the pressure flow hypothesis.
As glucose is prepared at the source (by photosynthesis) it is converted to sucrose (a disaccharide).
The sugar is then moved in the form of sucrose into the companion cells and then into the living phloem sieve tube cells by active transport.
This process of loading at the source produces a hypertonic condition in the phloem.
Water in the adjacent xylem moves into the phloem by osmosis.
As osmotic pressure builds up the phloem sap will move to areas of lower pressure.
At the sink osmotic pressure must be reduced.
Again active transport is necessary to move the sucrose out of the phloem sap and into the cells which will use the sugar – converting it into energy, starch, or cellulose.
As sugars are removed, the osmotic pressure decreases and water moves out of the phloem.
To summarise, the movement of sugars in the phloem begins at the source, where sugars are loaded (actively transported) into a sieve tube.
Loading of the phloem sets up a water potential gradient that facilitates mass movement in the phloem.
Phloem tissue is composed of sieve tube cells, which form long columns with holes in their end walls called sieve plates.
Cytoplasmic strands pass through the holes in the sieve plates, so forming continuous filaments.
As hydrostatic pressure in the sieve tube of the phloem increases, pressure flow begins, and the sap moves through the phloem.
Meanwhile, at the sink, incoming sugars are actively transported out of the phloem and removed as complex carbohydrates.
The loss of solute produces a high water potential in the phloem, and water passes out, returning eventually to the xylem.